| |
Главная
|
ДоменыПервичная структура доменов константных областей пяти классов тяжелых цепей показана на рисунке: 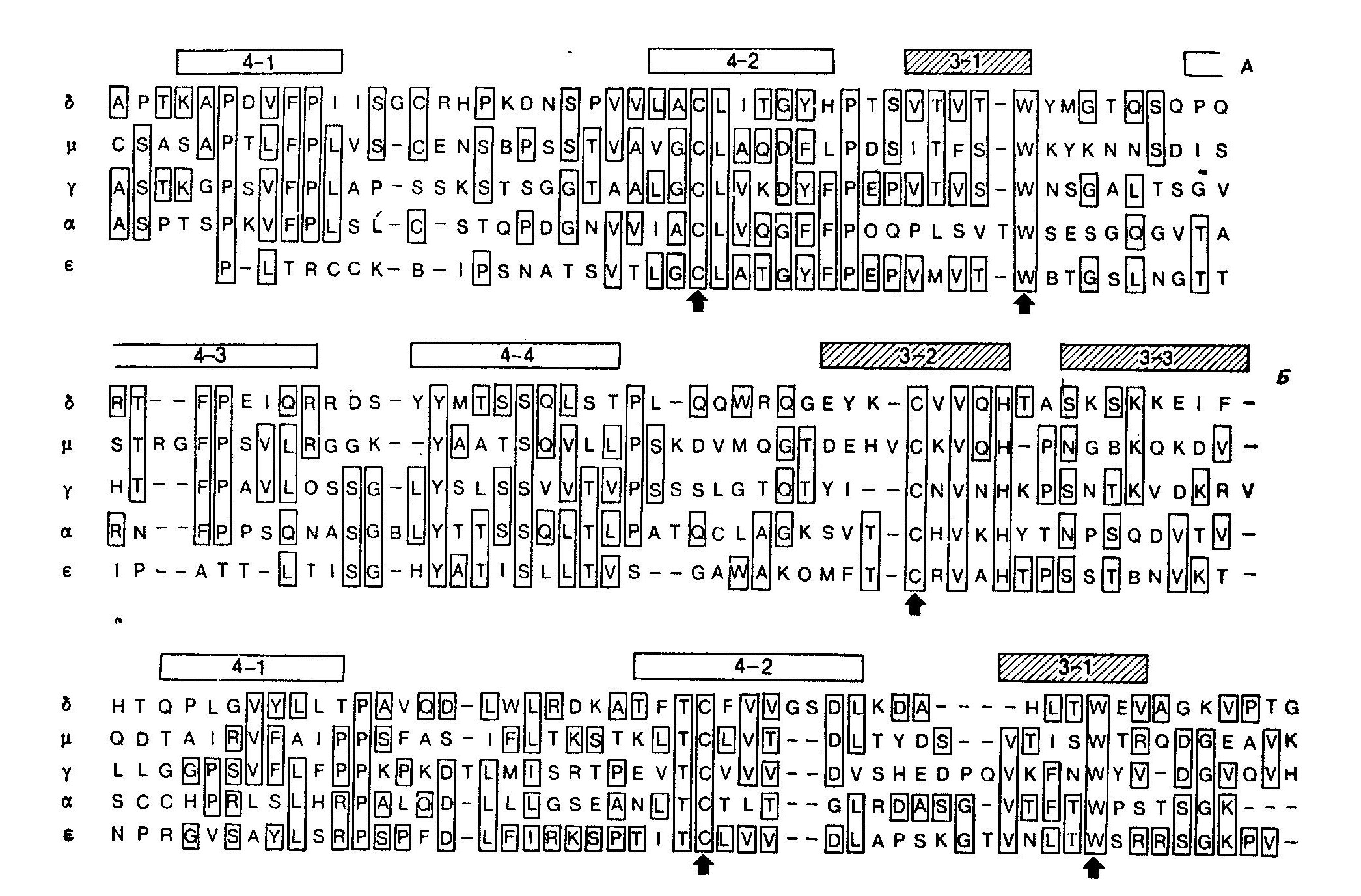  Пропуски введены для получения максимальной гомологии. Инвариантные остатки цистеина (С) и триптофана (W) в каждом домене отмечены стрелками. Последовательности аминокислот здесь расположены так, чтобы была получена максимальная гомология; гомологичные сегменты с бета-складчатостью обозначены согласно схеме Эдмундсона и др. Наибольшее внимание привлекают остатки, одинаковые для всех классов. В СН1 имеется 10 инвариантных положений, восемь — в первом домене Fc и девять — в карбоксиконцевом домене. Два цистеина, образующие дисульфидную связь, и триптофан, расположенный на расстоянии 14—16 остатков от первого цистеина, присутствуют всегда и, по-видимому, формируют ядро домена в процессе свертывания цепи. Другие малоизменчивые остатки расположены главным образом в сегментах, формирующих бета-складчатость. Замены в этих сегментах обычно наблюдаются между парами сходных по свойствам аминокислот. У сегментов, занимающих края бета-складчатых слоев, гомологичны лишь отдельные, разбросанные по цепи остатки. По-видимому, эти сегменты имеют менее важное значение для образования водородных связей, необходимых для поддержания стабильности домена. Большинство замен обнаружено в областях, расположенных вне бета-складчатых слоев, в частности в участках с беспорядочной конформацией. Степень гомологии каждого данного константного домена по отношению к любому другому константному домену белка того же вида равна примерно 25—30%. Это относится к постоянным областям как легких, так и тяжелых цепей. CHI-домены разных классов обладают наибольшей гомологией между собой, достигая 33%, что, возможно, отражает их общую функцию, заключающуюся в образовании пары доменов легкой цепи. Более всего схожи друг с другом Cγ1 и Сε1 (45% гомологии). У Fc-чаcтей молекул гомология составляет 30%. Однако при более подробном сравнении оказывается, что гамма- и эпсилон-цепи сходны почти на всем протяжении своих Fc-чаcтей, тогда как у мю- и альфа-цепей одни домены гомологичны лишь на 20%, а карбокси- концевые домены — на 50%. Это означает, что эволюция доменов происходила при разной скорости мутаций или же что гены, кодирующие отдельные домены, участвовали в процессе рекомбинации или конверсии. Гомология каждого из CH1-доменов при сравнении с любым СН2-, СНЗ- или СН4-доменом других четырех цепей составляет 25%. Скорее всего, каждый из постоянных доменов эволюционировал независимо; менее вероятно, что группа доменов одной тяжелой цепи является прямым предшественником другой тяжелой цепи. При сравнении константных областей одного класса иммуноглобулинов разных видов выявляется значительная эволюционная консервативность, поскольку гомология по большей части составляет примерно 60%. Отсюда следуют по крайней мере две возможности: а) все пять классов иммуноглобулинов существовали до образования известных видов млекопитающих, или же б) гены-предшественники эволюционировали у каждого вида в сторону выполнения сходных функций и в результате функциональных ограничений должны были привести к закреплению определенной структуры. Как уже указывалось, IgM менее всего изменчивы и обнаруживают 80%-ную гомологию при сравнении последовательностей тяжелых цепей человека, собаки и мыши. Очевидно, что ограничения, накладываемые функцией IgM, выражены более всего по сравнению с другими классами иммуноглобулинов. Гомология цепей подклассов одного класса иммуноглобулинов, как правило, составляет от 60 до 90% или даже более. Это указывает на сравнительно недавнюю дупликацию соответствующих генов у каждого вида. Очевидно также, что развитие подклассов шло независимо у разных видов. |