| |
Главная
|
Сайт-специфическая рекомбинацияПри сравнении нуклеотидных последовательностей клонов гаметных генов V и С можно точно указать сайт, в котором произошло соединение гаметных генов V и J1 в лимфоидных клетках, давших начало МОРС41. 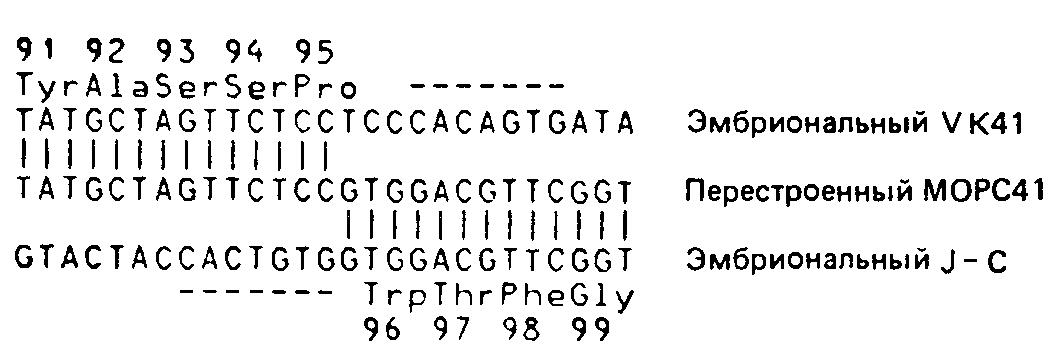 Последовательность перестроенного и экспрессирующегося гена МОРС41 приведена вместе с последовательностью гаметного предшественника F-области (эмбриональный VK41) Как видно из рисунке, перестроенный клон совпадает с гаметным клоном V до второго нуклеотида кодона 95; очевидно также, что перестроенная последовательность после этой точки происходит уже из Л. Всегда ли рекомбинация происходит точно между вторым и третьим нуклеотидами кодона 95? Анализ аминокислотных последовательностей х-цепей позволяет предположить, что, напротив, система рекомбинации обладает некоторой гибкостью. Выше уже было отмечено, что аминокислотные последовательности всех х-цепей с известной первичной структурой соответствуют тому или иному гаметному J-сегменту в области 97—108. Существуют, однако, х-цепи, у которых амино-кислота-96 не соответствует ни одному из гаметных J-фрагментов. Эти цепи могли бы кодироваться теми перестроенными генами, у которых соединение между V- и J-фрагментами произошло между разными нуклеотидами внутри кодона 96. 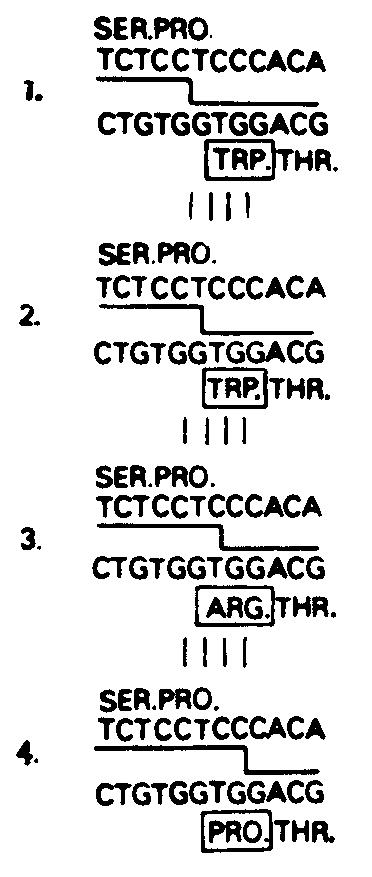 Первый пример иллюстрирует участок рекомбинации в МОРС41 — между вторым и третьим нуклеотидами кодона 95. Это наблюдение может объяснить необычайно высокую вариабельность аминокислот в положении 96. Сравнение перестроенной и гаметной форм других генов также позволяет предположить, что точки рекомбинации в них обладают некоторой подвижностью. |