| |
Главная
|
Механизм рекомбинацииВ поисках подхода к выяснению механизмов рекомбинации был проведен сравнительный анализ последовательностей гаметных генов F-области и гаметных генов J. 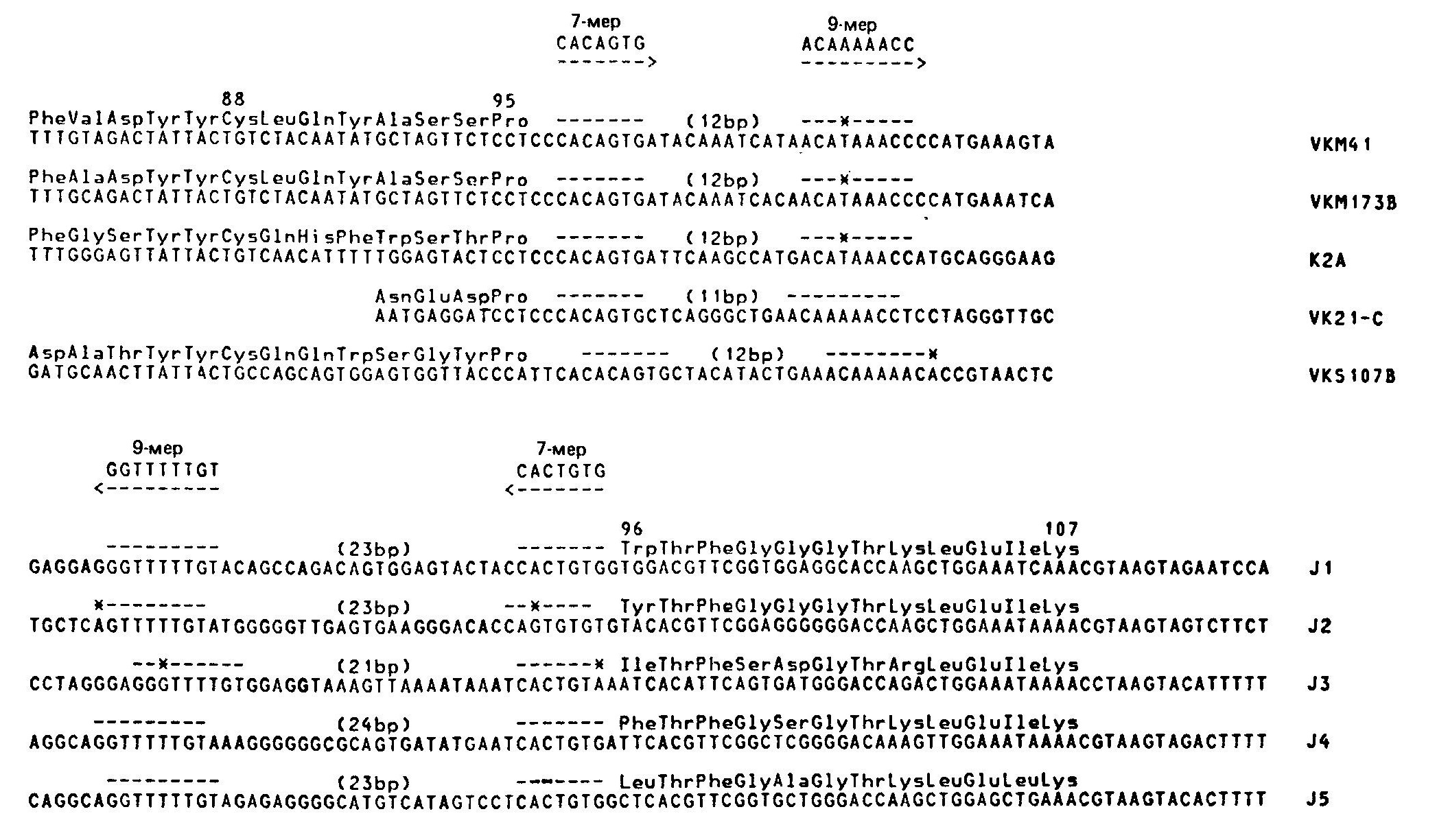 Несколько последовательностей гаметных генов Ух мыши и 5 последовательностей генов JX расставлены в соответствии с гомологией их структур. Единственно сопоставимые последовательности в генах V n J — это обозначенные прерывистыми линиями гептамеры и нонамеры (звездочками указаны встречающиеся отличия от «идеальных» последовательностей гептамера и нонамера). Два этих элемента разделены промежутком (спейсером) в 11—12 пар оснований в последовательности, соседствующей с F-областью, и спейсером в 23—24 пары в последовательности, соседствующей с J-областью. Были обнаружены два консервативных блока последовательностей, которые, возможно, играют роль в рекомбинации. Первый из них — это гептамер CACTGTG, расположенный на 5'-концах генов J, и комплементарный ему (инвертированный) гептамер CACAGTG на З'-концах генов F. Второй — нонамер GGTTTTTGT, расположенный на расстоянии 22 нуклеотидов от 5'-конца гептамера J и комплементарный ему АСАААААСС на расстоянии 11 нуклеотидов от З'-конца гептамера V. Важное значение этих элементов для рекомбинации подтверждается их наличием в сходных позициях генов К- и Н-цепей, а также отсутствием по обе стороны от этих генов других консервативных последовательностей. Комплементарность гептамеров и нонамеров, принадлежащих генам V и J, позволила предположить что эти элементы могут участвовать в образовании структуры «стебля и петли», являющейся промежуточной стадией рекомбинации. 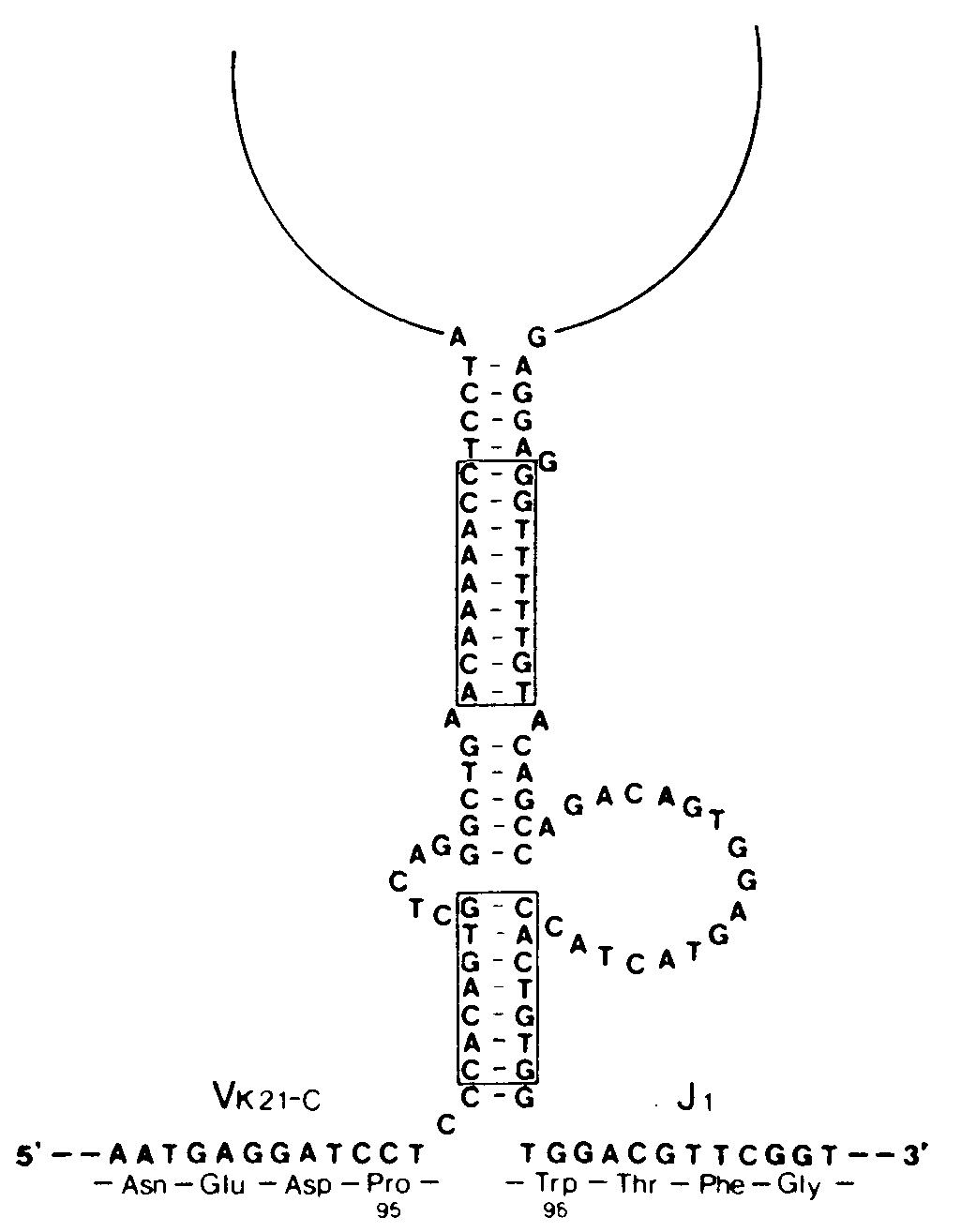 Этот предполагаемый промежуточный продукт рекомбинации V — J может возникать в результате комплементарности гептамерных и нонамерных элементов (обведены рамкой)', при образовании пар оснований между соответствующими элементами последовательностей V и J. Такая структура, по всей видимости, в свободном виде термодинамически невыгодна, однако она могла бы стабилизироваться специфическими белками. При образовании «стебля и петли» кодирующие последовательности генов V и J сближаются; при этом они либо соединяются под действием топоизомеразы I, либо транскрибируются, образуя непрерывную цепь ДНК, по механизму «выбора копии», когда при образовании новой цепи, комплементарной последовательностям V и J, пропускается участок ДНК', входящий в «стебель с петлей». С другой стороны, последовательности гептамера и нонамера могут узнаваться участвующими в рекомбинации белками; в этом случае необходимость в образовании комплементарных пар и структуры типа «стебель с петлей» отпадает. Одно из предсказаний модели «стебля и петли» состоит в том, что ДНК, образующая эту структуру, должна при рекомбинации вырезаться. Данные, полученные на нескольких миеломах, казалось, подтвердили это предположение, но более поздние результаты показали, что такое вырезание сопровождает рекомбинацию не всегда. В результате этих более поздних работ была предложена модель, основанная на неэквивалентном обмене между сестринскими хроматидами. В этой модели предполагается, что в течение части клеточного цикла (после репликации ДНК, но до деления клетки) /-область одной из хроматид реципрокно рекомбинирует с F- областью другой хроматиды путем взаимного обмена. При последующем делении клетки две новые хроматиды расходятся, давая начало дочерним клеткам двух типов. Один тип клеток будет содержать «нормальную» рекомбинацию V — J, описанную выше. Эти клетки будут выглядеть так, как будто вся ДНК между рекомбинировавшими F и J делетирована и результаты блоттинга по Саузерну будут подобны описанным ранее. Клетки другого типа будут содержать продукт реципрокной рекомбинации: со стороны 5'-конца от участка рекомбинации будут располагаться гептамер и нонамер из принимавшей участие в рекомбинации J-области, а со стороны 5'-конца от него появятся гептамер и нонамер, располагавшиеся до этого со стороны З'-конца от рекомбинировавшей F-области. 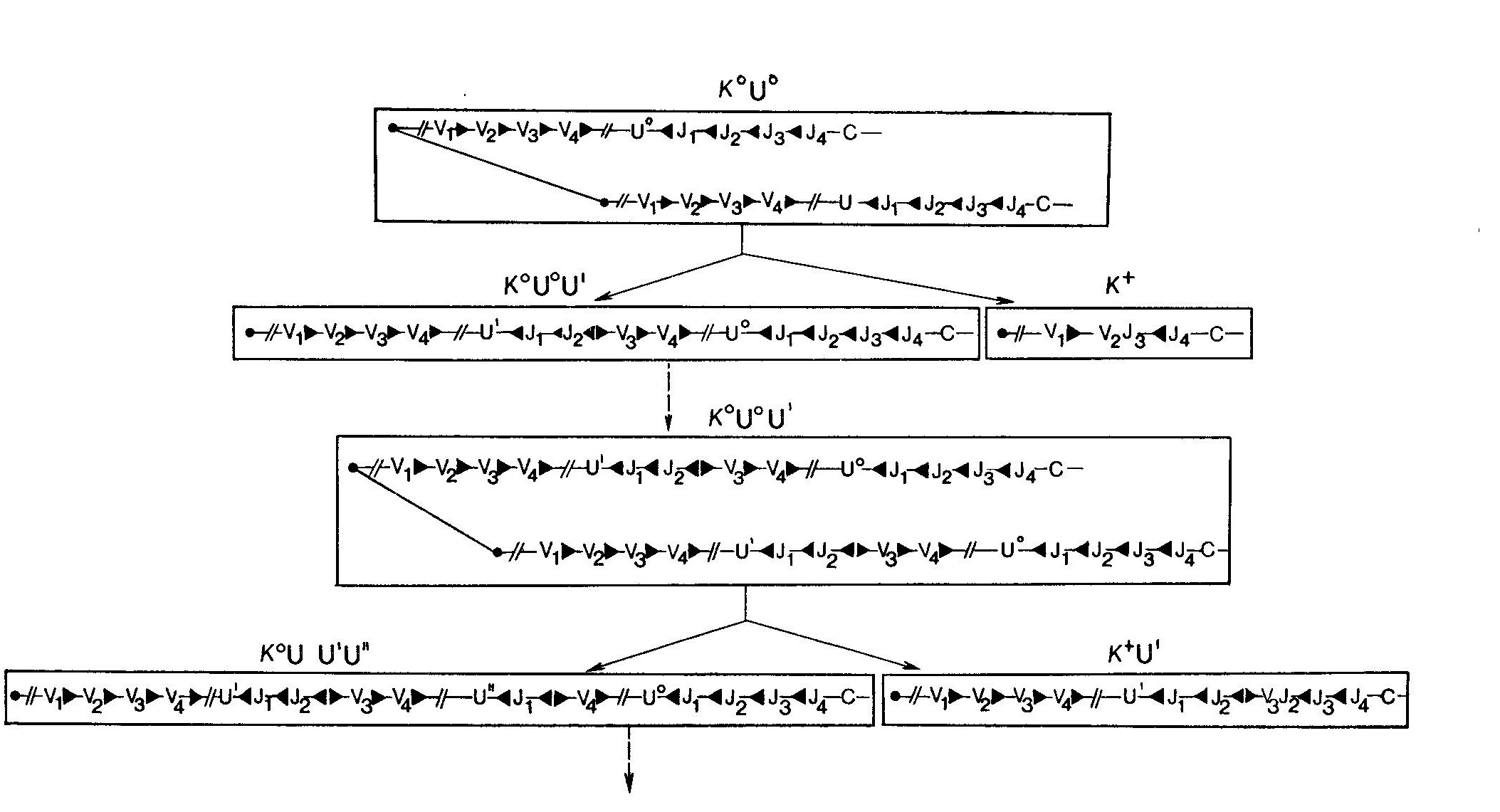 В каждом из прямоугольников изображен локус х одной хромосомы (отцовской или материнской). Прослежены события в течение двух клеточных делений. х° и х+ обозначают состояние гена Ск в геноме; ген х° — это гаметная конфигурация гена, а х+ — ген, подвергшийся функциональной перестройке V-J. «U» обозначает фрагмент ДНК, расположенный левее (с 5'-стороны) от фрагментов JX, который может существовать в гаметной конфигурации (U0) либо подвергаться различным перестройкам (т. е. U', U") вследствие рекомбинации в локусе J Черные треугольники — это гептамерный и нонамерный сигнальные элементы, примыкающие (фланкирующие) к V- и J-областям. В верхнем прямоугольнике располагаются две идентичные хроматиды в гаметной конфигурации (К0, U0), готовые для НОСХ между V2 и J3. Прямоугольники под ним иллюстрируют строение двух продуктов НОСХ, расходящихся по двум дочерним клеткам. В правом прямоугольнике — продукт функциональной перестройки (х+) между V2 и J3 с кажущейся делецией фрагмента ДНК, располагавшегося между ними в гаметной конфигурации. В левом прямоугольнике — этот же фрагмент, дуплицированный во втором продукте обмена. Этот продукт сохраняет гаметные последовательности К0 и U0 и приобретает (вместе с дуплицированным фрагментом) вторую, перестроенную 5'-последовательность (U'). Содержащий продукты НОСХ локус х дочерней клетки может подвергнуться НОСХ вторично (прямоугольники внизу); при этом могут образоваться К+-клетки, сохраняющие перестроенную 5'-последовательность (K+U', прямоугольник справа). Черные треугольники, расположенные навстречу друг другу, представляют продукты рекомбинации по фланкирующим маркерам. В этих клетках последовательность между участвовавшими в рекомбинации F и J, т. е. участок ДНК, удаляемый в процессе «нормальной» рекомбинации F — J, будет дуплицирован. Кроме того, в этих клетках будет содержаться полный набор генов F и J, которые при дальнейшей рекомбинации могут дать начало совершенно правильным рекомбинантам V — J. Поскольку рекомбинация эта происходит в пределах одной из дуплицированных хромосом (а не между гомологичными материнской и отцовской хромосомами), другая хромосома может при этом остаться в гаметной конфигурации либо принять участие в другой, независимой последовательности аналогичных перестроек. Во всех случаях ни на одной из стадий ген С не дуплицируется. Поэтому при блоттинге по Саузерну геномной ДНК можно обнаружить не более двух гомологичных копий (принадлежащих отцовской и материнской хромосомам) вне зависимости от числа произошедших в данной клеточной линии обменов между сестринскими хроматидами. Эта модель позволяет сделать важное предсказание, не укладывающееся в простую модель «делеции петли»: часть клеток должна содержать продукты реципрокной рекомбинации с прилежащими друг к другу гептамерами F и J и фланкирующими (расположенными по обе стороны) нонамерами. Это предсказание подтвердили Хехтль и др., сообщившие о клонировании и секвенировании таких фрагментов (названных авторами продуктами «фланкирующей» рекомбинации) из трех различных миелом.Ни в одном из этих трех случаев продукт рекомбинации, включающий фланкирующие последовательности, не был обнаружен в клетке, содержащей нормальный V — J-продукт. Именно такой результат и был предсказан моделью, так как продукты взаимной рекомбинации всегда должны расходиться по разным дочерним клеткам. Во всех трех случаях фрагмент, происходящий от J-области, принадлежал гену J1. Фрагменты, происходящие от F-области, не соответствовали ни одной из опубликованных к тому времени последовательностей гаметных F-областей, но, используя в качестве зонда происходящий от гена V фрагмент из З'-области одного из фланкирующих продуктов, Хехтль и др. смогли проклонировать соответствующий ему гаметный ген V. Оказалось, что все три продукта, содержащие фланкирующие последовательности, обладают интересным свойством: оба гептамер а прилегают друг к другу так плотно, что между ними нет ни одного промежуточного нуклеотида. Для получения правильных реципрокных продуктов при перестройках V-J в МОРС41, МОРС173В и S107B, согласно предсказанию, должны были бы существовать несколько промежуточных нуклеотидов. Это, видимо, необходимо для всех перестроек с сохранением рамки считывания, так как гептамерная последовательность отделена от кодона 95 га-метного гена V и от кодона 96 гена J несколькими нуклеотидами. В процессе перестройки, возможно, происходит делеция этих нуклеотидов. Модель неравного обмена между сестринскими хроматидами подтверждена и другим исследованием, показавшим, что клетки, несущие продукты рекомбинации, содержащие фланкирующие последовательности, встречаются достаточно часто. Ван Несс и др. изучили 28 миелом с перестроенными генами х и выяснили, что в 15 из них при гибридизации по Саузерну с фрагментом ДНК, расположенным со стороны 5'-конца от генов J-области, обнаруживаются полосы, соответствующие, вероятно, фланкирующим фрагментам. Не обусловлено ли образование про-дуктов рекомбинации, содержащих фланкирующие последовательности, аномальным перестройками, связанными с нефизиологическими условиями в миеломной ткани? Для ответа на этот вопрос Ван Несс и др. использовали метод количественного определения интенсивностей полос, получаемых при блоттинге по Саузерну ДНК из отобранных при помощи проточной цитофлуориметрии х-синтезирующих В-лимфоцитов (как описано в предыдущем разделе). 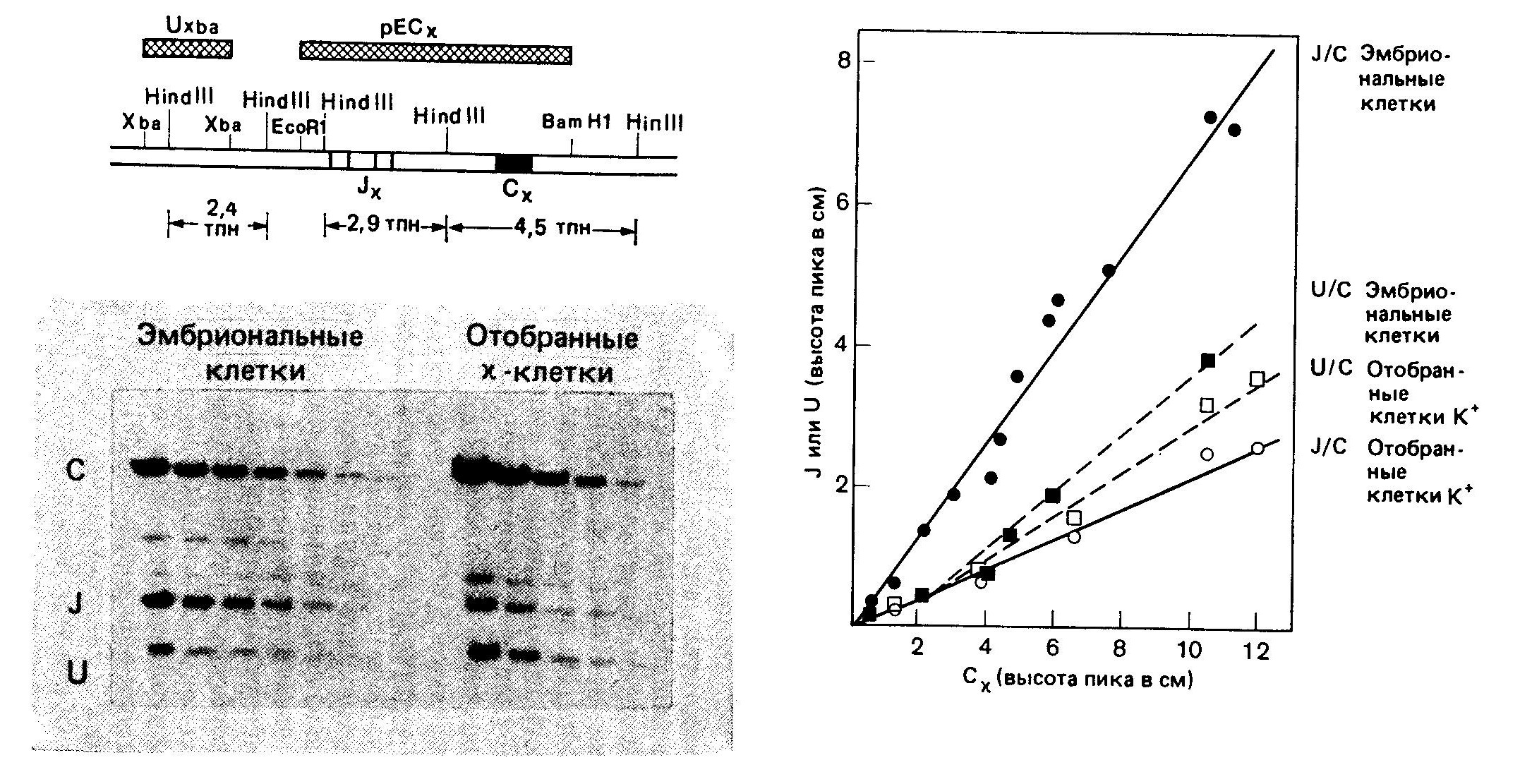 На картинах блоттинга по Саузерну расщепленной Hind III ДНК из мышиных эмбрионов (гаметной ДНК) и из х+-лимфоцитов (отобранных с помощью проточного цитофлуориметра) зонды (слева вверху) выявляют главные полосы длиной 2,4; 2,9 и 4,5 тпн, представляющие соответственно «левые» 5'-концевые последовательности, /-фрагменты и область Сх. Интенсивность полосы Су (4,5 тпн), не зависящая от генных перестроек, использовалась для нормализации интенсивностей других полос. Для нахождения линейной области ДНК наносилась на гель в нескольких концентрациях. Определив отношение нормализованных интенсивностей графиков //С (для отобранных х+-клеток) к //С (для эмбрионов), Ван Несс и др. рассчитали, что х+-клетки сохраняют около 30% последовательностей локуса J, содержавшихся в гаметной ДНК. Отношение U/C (х+-клетки) к U/C (эмбрионы) указывало, что около 75% «левых» 5'-концевых последовательностей также сохранялось. Эти результаты согласуются с моделью НОСХ и противоречат модели с делецией петли. Выбор участков расщепления рестриктазами и гибридизационного зонда позволил независимо определить интенсивность полосы гена С (ни интенсивность, ни размер которой не должны изменяться при перестройках V — J), гаметной полосы J (интенсивность которой должна падать при пере-стройке) и полосы, представляющей последовательность, располагающуюся со стороны 5'-конца от J-области (интенсивность которой может уменьшаться только при делециях этой последо-вательности из генома). Результаты по-казали, что примерно две трети локусов х содержат перестроенные гены J (как описано в предыдущем разделе), но удалена лишь четверть всех 5'-последовательностей. Отсюда следует, что в ДНК из популяции лимфоцитов большая часть перестроек не сопровождается делециями последовательностей, расположенных со стороны 5'-конца от генов /. Это со-гласуется с моделью обмена между сестринскими хроматидами. Хотя, как следует из вышесказанного, механизм рекомбинации V — / остается во многом непонятным, некоторые обобщения и выводы уже можно сделать. Консервативные гептамер и нонамер остаются кандидатами на роль участков, узнаваемых при рекомбинации. Основанная на этих элементах структура «стебель и петля» — также вполне вероятный промежуточный этап рекомбинации, но основанная на делеции этой структуры модель сейчас кажется менее убедительной, чем в то время, когда она была предложена. В соответствии с этим и сама структура стала казаться несколько менее привлекательной. Модель обмена между сестринскими хроматидами, видимо, согласуется со всеми имеющимися данными. Клонированные Хехтлем и др. фланкирующие фрагменты прекрасно согласуются с этой моделью, однако то, что они являются продуктами взаимной рекомбинации при функциональных (в отличие от ошибочных) перестройках V — J, не было однозначно показано. Чтобы доказать, что функциональная перестройка происходит при обмене между сестринскими хроматидами, нужно было бы сравнить все четыре последовательности, участвующие в одном рекомбинацйонном событии,— двух родительских генов (V и С) и двух перестроенных генов (функционального рекомбинанта VJ и рекомбинанта, содержащего фланкирующие участки), — однако такой набор клонов будет, по всей видимости, нелегко создать. Точный ответ вероятно, можно получить в опытах на системах рекомбинации in vitro; такие системы разрабатываются в нескольких лабораториях. Несмотря на то что некоторые детали механизма рекомбинации пока не выяснены, общая модель, основанная на изучении генов х, получила широкое признание. Согласно этой модели, гаметный геном содержит множество генов F^-области (возможно, несколько сотен, см. ниже) и одну С-область с 5 (4 актив-ными) J-генами на ее 5'-конце. На некотором этапе развития В-лимфоцита один из генов V рекомбинирует с одним из активных генов J. Если такая перестройка функциональна, рекомбинация коммитирует клетку и все ее потомство к экспрессии этой перестроенной F-области. |