| |
Главная
|
Локус аВскоре после того как Удин идентифицировал в IgG три основных аллотипа локуса a (a1, а2 и а3), а Тодд вслед за этим показал, что эти аллотипы встречаются и у иммуноглобулинов других классов, стало ясно, что аллотипические детерминанты, кодируемые локусом а, находятся в VH -областях, которые могут присоединяться к СH-области любого изотипа. Поскольку в гаметной ДНК, по-видимому, имеется множество генов VH, аллотипические замены аминокислот, характерные для каждого аллеля (a1, а2 или а3), очевидно, должны повторяться во многих гаметных VH -генах одной хромосомы. Возникает парадокс, каким образом такой обширный набор генов VH может наследоваться в качестве «аллелей» и при этом на затрагиваться рекомбинациями. Одно из возможных объяснений состоит в том, что гены VH всех трех аллелей локуса а (a1, а2 и а3) находятся в одной хромосоме, а аллелизм обусловливается расположенным по соседству регуляторным геном. 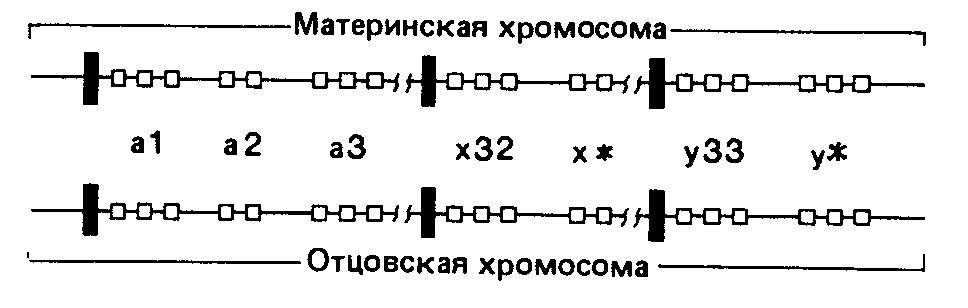 Группы генов VH (светлые квадраты), соответствующие определенным VH-аллотипам (а1,
а2, а3 и т. д.), Альтернативное объяснение предполагает возможность коррекции ограниченного числа рекомбинаций среди многочисленных аллельных генов VH путем конверсии генов. Молекулы аллотипов а1, а2 и а3 различаются между собой заменой по меньшей мере шести аминокислот N-концевых каркасных участков VH-области, а возможно, и дополнительными заменами в других участках VH. Это позволяет отнести их к комплексным аллотипам, т. е. имеющим не одну-две аминокислотные замены, как следует ожидать для простых аллельных вариантов, а многочисленные замены. Результатом этого является наличие в молекуле нескольких аллотипических детерминант, которые обусловливают возможность преципитации иммуноглобулинов а-аллотипа антиаллотипической антисывороткой. Встречаются и латентные аллотипы локуса а. В некоторых случаях они неожиданно экспрессируются у кроликов, для которых предсказать подобную экспрессию, исходя из их генетической характеристики, было невозможно. В классической работе Стросберга и др.было обнаружено, что гетерозиготный кролик a1a3 отвечал на иммунизацию бактериальными клетками образованием иммуноглобулинов с а2-аллотипом наряду с a1- и а3-иммуноглобулинами. Этот факт находится в явном противоречии с законом Менделя, согласно кото-рому у( одной особи может присутствовать не более двух аллелей одного гена. Аналогичное явление наблюдалось и для локуса Ъ легких х-цепей: у того же самого гетерозиготного кролика b4b5 обнаруживались и х-цепи b6. Подобные неожиданные латентные аллотипы локусов a, b и других были найдены (хотя и в очень малых количествах) также у некоторых неиммунных кроликов. Такое парадоксальное поведение аллотипов локуса а более или менее хорошо объясняется тем, что они представляют собой три сцепленные группы гомологичных гаметных структурных генов VH, имеющих отношение к каркасным участкам и присутствующих у всех кроликов. Таким образом, каждая VH- группа (a1, а2 или а3) — это псевдоаллель, а настоящий аллелизм присущ регуляторному гену, контролирующему экспрессию генов VH. Данное предположение позволяет также объяснить экспрессию аллотипов локуса b (Сх) и, возможно, локусов х, у и d. Значительная часть (10—30%) тяжелых цепей иммуноглобулинов каждого кролика не имеет ни a1-, ни а2-, ни а3-аллотипических детерминант; такие цепи называют «а-отрицательными». Относительное содержание а-отрицательных молекул можно увеличить с помощью супрессии, подавляя, например, биосинтез а2-детерминанты у гомозиготного кролика а2/а2. Получающаяся в результате популяция иммуноглобулинов оказывается полностью а-отрицательной. При иммунизации такими иммуноглобулинами гомозиготного кролика a1/a1 были выявлены еще два неаллельных аллотипа, х32 и уЗЗ. Таким образом, х и у представляют собой два дополнительных локуса, причем они тесно сцеплены с локусом а. Локус х представлен двумя аллелями: х и еще одним аллелем, детерминанта которого не выявлена. Аналогичным образом локус у соответствует детерминанте уЗЗ и ее неиммуногенному аллельному варианту. Аллотипические детерминанты х и у (так же как и детерминанта а) находятся в VH-области. Следовательно, а, х и у — это по меньшей мере три тандемно сцепленные группы VH-генов; экспрессия каждой группы может контролироваться аллельными регуляторными генами. |