| |||||||||||
Главная
|
Строение лимфоидных органов и иммунные реакцииФункциональное значение микроокружения, существующего в лимфоидных органах, для развития иммунных ответов in vivo было наглядно продемонстрировано в экспериментах по изучению процессинга антигена с помощью элементов стромы и временных взаимоотношений между этим процессингом и активацией лимфоцитов. Как правило, антигены поступают в лимфатические узлы по приносящим лимфатическим протокам, впадающим в субкапсулярные синусы, а в селезенку по кровеносным сосудам, омывающим маргинальный синус. В лимфоидных органах обоих типов эти места обогащены макрофагами, быстро фагоцитирующими введенный материал. Большая часть такого захваченного антигена, по-видимому, разрушается, не внося сколько-нибудь существенного вклада в развитие иммунного ответа. Однако часть антигена, вероятно, остается в ассоциированном с поверхностью клеток виде и через 18—24 ч после иммунизации появляется в весьма специфическом виде внутри первичных фолликулов. Хотя какая-то доля этого фолликулярного антигена может быть связана с лимфоцитами, большая его часть, несомненно, локализуется на поверхности отростков фолликулярных дендритных клеток. До сих пор непонятно, каким образом эти клетки удерживают антиген; предполагается, что это происходит при прямом или непрямом участии предсуществующих специфических антител. Класс антител и их способность фиксировать комплемент имеют, по-видимому, важное значение для фолликулярной локализации комплексов антиген — антитело. Согласно одной из гипотез, фолликулярные дендритные клетки экспрессируют поверхностные рецепторы для Fc- областей иммуноглобулинов или для компонентов системы комплемента и благодаря этим рецепторам избирательно взаимодействуют с антителами, связанными с антигеном. Однако возникает вопрос об изначальном источнике антиген- специфических антител, поскольку многие антигены обнаруживаются в фолликулах еще до того, как появляется заметное количество сывороточных антител или специфических антителообразующих клеток. Фолликулярную локализацию можно наблюдать даже после облучения животных такими дозами, которые вызывают гибель практически всех В-лимфоцитов. Не исключено, что антителами, опосредующими связывание антигенов с фолликулярными дендритными клетками, служат какие-то перекрестно реагирующие или «естественные» антитела, уже присутствующие в сыворотке крови в низкой концентрации. Возможно, однако, что этот эффект достигается в результате взаимодействия ФДК с антиген-специфическими факторами, секретируемыми Т-клетками. Первое, что происходит в лимфоидном компартменте лимфатического узла в ответ на введение какого-либо сложного антигена, например бараньих эритроцитов (БЭ), — это волна процессов активации и деления лимфоцитов, возникающая через 1—2 дня после иммунизации. Частота Т-клеточных митозов становится максимальной приблизительно на 3-й день, а затем довольно быстро снижается, достигая обычного уровня на 7-й или 8-й день. Частота В-клеточных митозов доходит до максимума примерно на один день позже и остается сравнительно высокой в течение более продолжительного времени. Антителообразующие клетки, секретирующие антитела против бараньих эритроцитов, преимущественно IgM-класса, появляются на 3-й или 4-й день и скоро оказываются основным компонентом мякотных тяжей. Начиная с 4-го или 5-го дня большинство делящихся В-клеток собирается в обособленные группы, называемые зародышевыми центрами, которые связываясь со специфическим антигеном, располагаются на поверхности фолликулярных дендритных клеток. Маловероятно, чтобы зародышевые центры участвовали в генерировании первичного иммунного ответа, поскольку их удается достоверно идентифицировать только после первого появления сывороточных антител. Аналогичное поведение В- и Т-клеток наблюдается в селезенке после внутривенного введения бараньих эритроцитов. Время протекания всех этих реакций, выявленных гистологическими методами, равно как и их относительный вклад в иммунный ответ, сильно варьируют и зависят в значительной степени от природы иммунизирующего антигена. Так, например, более ярко выраженная и более длительная реакция, происхо-дящая в паракортикальном веществе, является, вероятно, основным проявлением ответов на антигены, вызывающие Т-клеточный иммунитет (к таким антигенам относятся оксазолон и другие контактные сенсибилизаторы или аллотрансплантаты). При стимуляции некоторыми полимерными антигенами, индуцирующими Т-независимый гуморальный ответ (преимущественно антитела IgM- и IgG3-MaccoB) образование плазматических клеток происходит на фоне относительно слабой паракортикальной Т-клеточной реакции, не сопровождаясь появлением отчетливо выраженных зародышевых центров. С другой стороны, развитие Т-зависимого гуморального ответа сопровождается активным формированием зародышевых центров, что, по всей видимости, служит необходимым условием эффективного образования антител IgG- и IgA- классов. Весьма важное значение имеет также и форма поступающего антигенного материала. Корпускулярные и агрегированные антигены, а также комплексы антиген-антитело, образованные при избытке антигена, вызывают оптимальный ответ, тогда как те же самые антигены в растворимой форме часто являются слабыми иммуногенами, что проявляется в возникновении замедленной и слабой ответной реакции. Лимфобласты, образующие зародышевые центры, обладают уникальным фенотипом: помимо своих больших размеров (в среднем), что является следствием их активированного состояния, эти клетки (у мышей и у людей) характеризуются высоким содержанием поверхностных участков связывания для PNA, растительного лектина, специфически взаимодействующего с концевыми остатками галактозы. 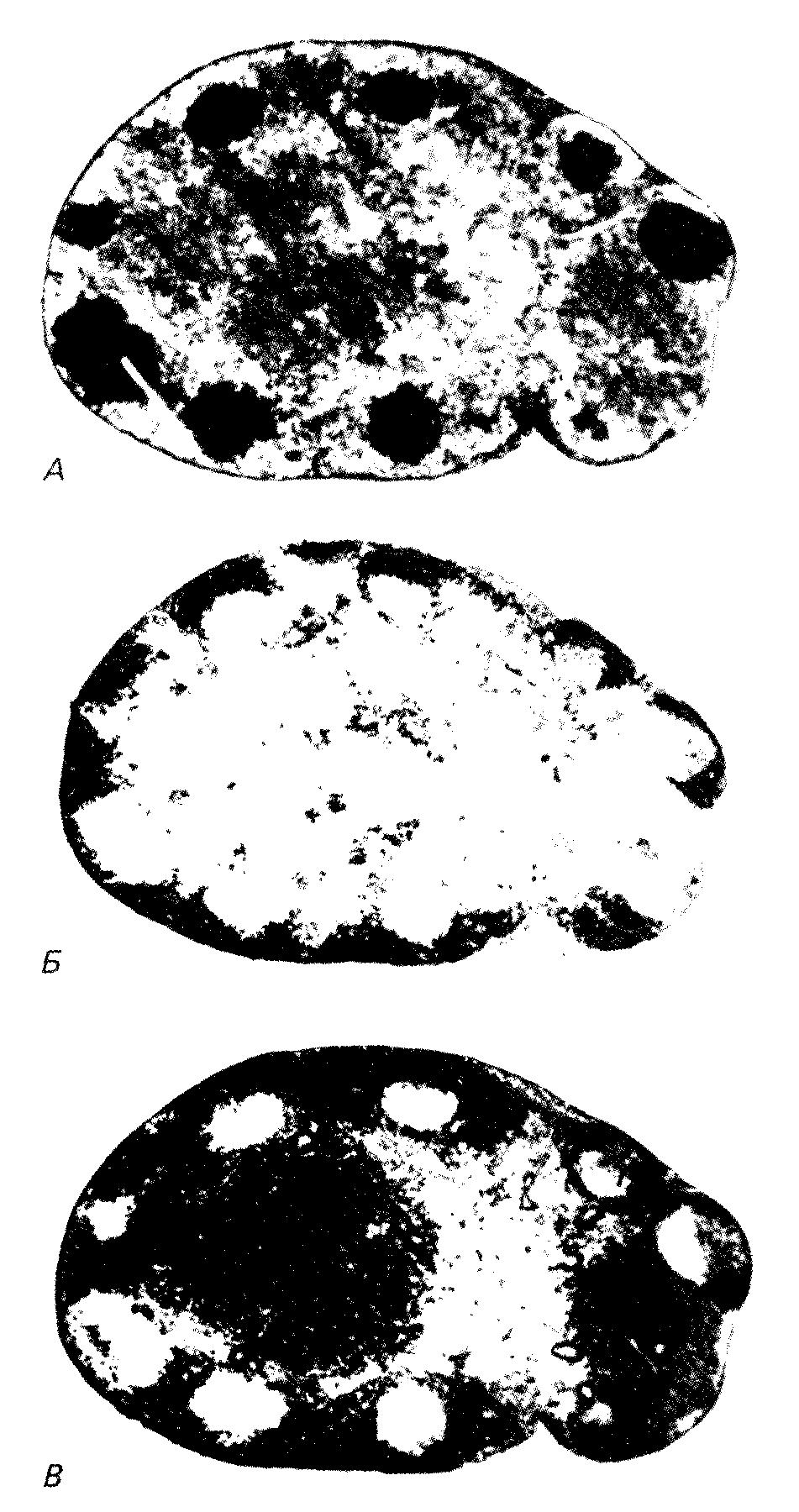 Через 7 дней после иммунизации бараньими эритроцитами удалили плечевой лимфатический узел. Это свойство позволяет отличать клетки зародышевых центров от других В-клеток, а также от плазматических клеток и от большинства плазмобластов. Ранее мы уже высказывали предположение о том, что PNA-связывающие участки могут либо быть ассоциированы с молекулами, помогающими удерживать дифференцирующиеся лимфоидные популяции в определенных местах, либо сами выступать в роли таких молекул, поскольку основные популяции лимфоцитов с фенотипом РКАbi-клетки зародышевых центров, тимоциты и, по-видимому, пре-В-клетки костного мозга — все представляют собой немигрирующие популяции, на дифференцировку которых оказывает влияние локальное микроокружение. Проведенные недавно эксперименты с использованием PNA в качестве маркера для идентификации В-клеток зародышевых центров в суспензиях стимулированных лимфатических узлов или пейеровых бляшек показали, что эти клетки находятся на одной из ключевых стадий В-клеточной дифференцировки. Возможно, самая существенная фенотипическая особенность клеток зародышевых центров — это экспрессия поверхностных Ig-рецепторов, специфичных по отношению к индуцирующему антигену. Действительно, после первичной стимуляции большинство антигенсвязывающих В-клеток обнаруживается в составе зародышевых центров лимфоидного органа. Эти клетки собираются в зародышевых центрах, по-видимому, благодаря своей способности взаимодействовать с антигеном, локализованным на поверхности ФДК, а затем под воздействием комплексов ФДК — антиген и Т-хелперов (и их продуктов) происходит образование клонов. Клетки зародышевых центров экспрессируют также уникальный набор дифференцировочных антигенов. В отличие от практически всех остальных фолликулярных лимфоцитов эти клетки лишены поверхностных IgD и в значительной степени дифференцировочного антигена ThB. Клетки зародышевых центров не мигрируют и не узнают высокоэндотелиальные венулы (ВЭВ), и соответственно этому у них отсутствуют предполагаемые рецепторы для ВЭВ, т. е. не наблюдается взаимодействия с моноклональными антителами MEL-14. По сравнению с IgD-B-клетками клетки зародышевых центров несут больше поверхностных антигенов, кодируемых I-A-областью, и меньше Н2К-детерминант; для них характерен также низкий уровень экспрессии поверхностных иммуноглобулинов (в 5—10 раз меньше, чем у окружающих малых В-лимфоцитов). 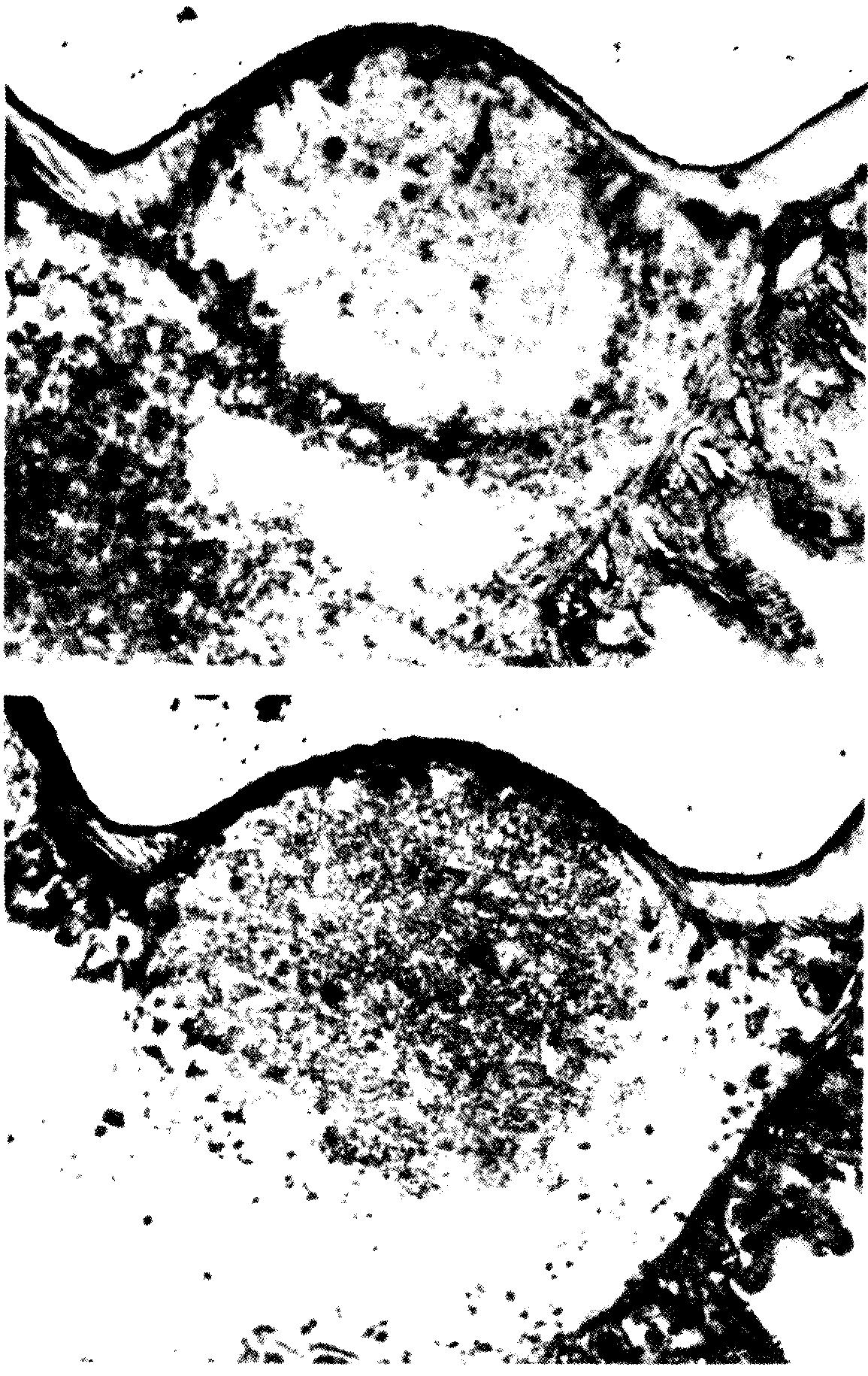 Вверху: окрашивание пейеровой бляшки конъюгатом кроличьих антител против Ig мыши с пероксидазой хрена. После первичной антигенной стимуляции основную часть этих поверхностных иммуноглобулинов составляют молекулы IgM, а вторичная или длительная стимуляция с помощью специфического антигена приводит к повышению содержания поверхностных IgG (в лимфатических узлах) или поверхностных IgA. Изотипы поверхностных иммуноглобулинов клеток зародышевых центров
Исходя из наблюдаемых изменений экспрессии класса тяжелых цепей, можно предположить, что помимо обеспечения антиген-специфических В-клеток определенным стимулом, необходимым для их клональной пролиферации, микроокружение внутри зародышевых центров играет важную роль в феномене переключения класса тяжелых цепей при развитии В-клеточного ответа. По-видимому, это осуществляется либо индукцией экспрессии, либо отбором соответствующих изотипов. Напрашивается вывод, что микроокружение в центрах размножения может оказывать влияние также и на другие стороны дифференцировки В-клеток. Например, зародышевые центры, расположенные в пейеровых бляшках, вероятно, программируют активированные В-клетки к экспрессии поверхностных рецепторов, специфичных по отношению к слизистому эндотелию, тогда как зародышевые центры лимфатических узлов, возможно, индуцируют экспрессию рецепторов для неслизистых детерминант (или регулируют отбор соответствующих клеток); в результате клетки впоследствии локализуются в тканях, сходных с теми, где произошла первоначальная антигенная стимуляция. Не исключено также, что зародышевые центры непосредственно участвуют в процессах, связанных с изменением аффинности антител, продуцируемых на разных стадиях В-клеточного ответа. При длительной стимуляции с помощью какого-либо антигена средний аффинитет антител к этому антигену постепенно увеличивается. Чтобы объяснить это явление, было высказано предположение, что это увеличение аффинности обусловлено конкурентным взаимодействием пролиферирующих клеток зародышевых центров с антигеном, находящимся на отростках фолликулярных дендритных клеток. Клетки, обладающие наиболее высоким аффинитетом или авидностью, избирательно стимулируются к пролиферации, в то время как клетки, экспрессирующие низкоаффинные антитела, не получают соответствующего сигнала и скорее всего погибают прямо в зародышевом центре. Согласно этой модели, микроокружение внутри зародышевых центров могло бы даже индуцировать или по крайней мере способствовать отбору В-клеток с соматическими мутациями, вызывающими образование антител с «улучшенной» специфичностью по отношению к иммунизирующему антигену. Ряд имеющихся данных указывает на то, что некоторые из перечисленных процессов отбора или отбраковки действительно происходят в зародышевых центрах. Во-первых (как и в случае дифференцировки пре-В-клеток и тимоцитов), скорость клеточной пролиферации намного превышает число идентифицируемых клеток, покидающих зародышевые центры. Во-вторых, результаты разнообразных гистологических экспериментов говорят о множественной гибели клеток — зародышевые центры содержат многочисленное множество аномальных («tingible body») макрофагов с фагоцитированными обломками ядер. Очевидно, что клетки зародышевых центров находятся на ключевой стадии В-клеточной дифференцировки. Однако пока что об их судьбе нам известно на редкость мало. По крайней мере некоторые из них, по-видимому, превращаются в дальнейшем в плазматические клетки, секретирующие иммуноглобулины. Другие, вероятно, дают начало В-клеткам памяти. Последняя возможность подтверждается широким спектром экспериментальных данных, демонстрирующих наличие корреляции между появлением зародышевых центров с одной стороны, и В-клеточной памяти — с другой. Дальнейшие исследования прямых предшественников клеток зародышевых центров, а также изучение судьбы клеток зародышевых центров в опытах с адоптивным переносом их реципиентам точнее определят роль микроокружения внутри зародышевых центров в возникновении и регуляции гуморального иммунитета. | ||||||||||